ひまわりの種を撒くと、わずか数センチの芽生えが、2カ月ほどもすれば人ほどの背丈にまで成長してしまう。たった数センチの細い茎が2メートルもの背丈を支え、数ミリの厚みの葉が広々と展開される。我々動物と同じ多細胞生物でありながら、植物はどのような仕組みでこうも違ったからだをつくり上げているのだろうか。
植物の体は細胞壁で包まれた細胞が石垣のように積み重なってつくられている。細胞自体がしっかりとかたちを保てなければ体をつくることはできない。植物の細胞壁は厚さわずか0.1マイクロメートルほどの薄い層であり、主成分であるセルロース微繊維と、それらを架橋する無数の糖鎖で構成される。分裂組織で生み出された細胞は水を吸って細胞壁を押し広げるように成長してゆく。しかし細胞壁に編み込まれたセルロース微繊維が「たが」となって細胞の膨張を制限するため、円柱状に伸長する。セルロース微繊維がうまく並んでいないと細胞は水風船のように球状に膨らんでしまう(図1)。植物の体は細長い細胞だけでなく、多様なかたちの細胞でつくられている。それぞれの細胞は、細胞壁の繊維をつくるタイミングや位置、方向を制御することでその場に適したかたちに成長する。「細胞壁のつくり方」が植物のからだづくりの鍵なのだ。
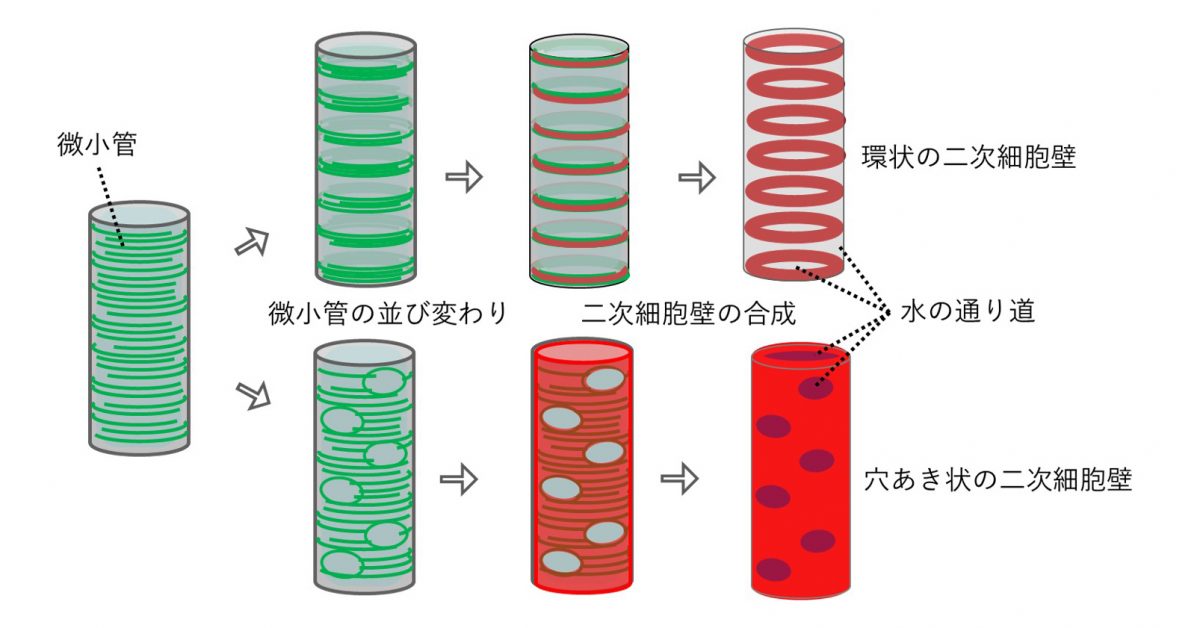
細胞壁の役割は細胞のかたちだけではない。木部など一部の組織では、成長した細胞が二次細胞壁とよばれる厚い細胞壁を発達させることにより、植物の体を支える丈夫な組織をつくり上げる。二次細胞壁がつくられる位置や方向も厳密に制御されている。その顕著な例が道管である。道管は、根から吸収した水分を地上の茎や葉に輸送する管状の組織である。もし道管が無ければ、植物はひまわりのように地上に高く体を成長させることはできない。道管の細胞は、二次細胞壁をつくった後、細胞壁以外の細胞内容物を自ら消化して管状の構造になる。二次細胞壁は疎水的で水を通さない性質を持つが、道管の二次細胞壁は細胞全体ではなく、環状や孔あき状につくられるため、水の通り道も確保される(図2)。「細胞壁のつくり方」が植物の体づくりの中でいかに重要かおわかりいただけただろう。
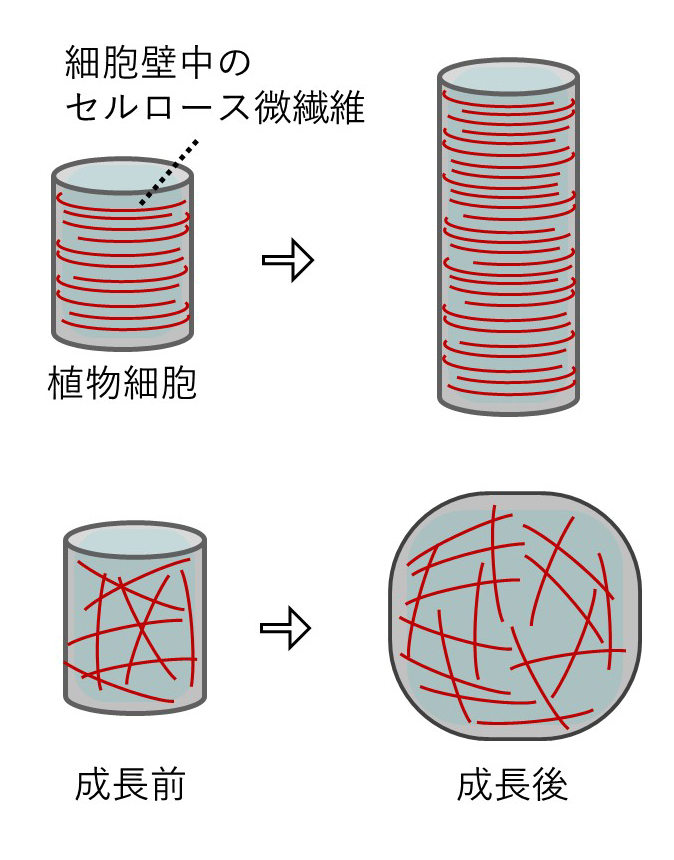
図1 細胞壁による細胞の成長制御
セルロース微繊維が並んでいると、その垂直方向に細胞が伸張するように成長する(上)。セルロース微繊維がうまく並んでいなければ球状に膨らんでしまう(下)。
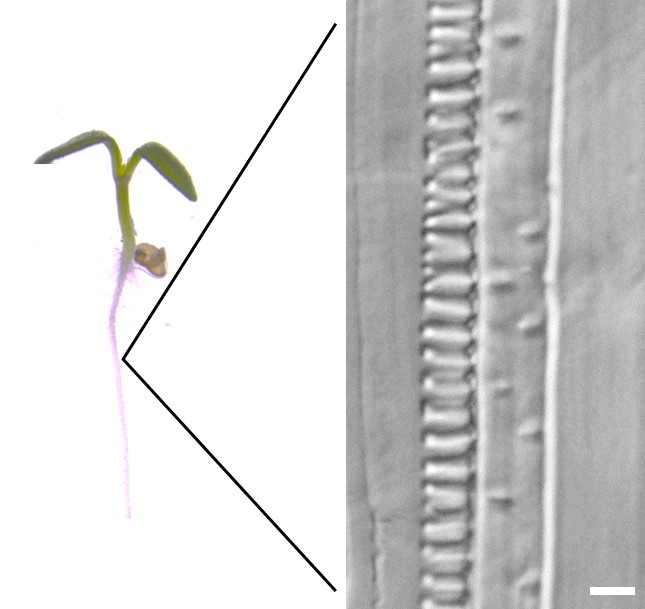
図2 道管の細胞壁
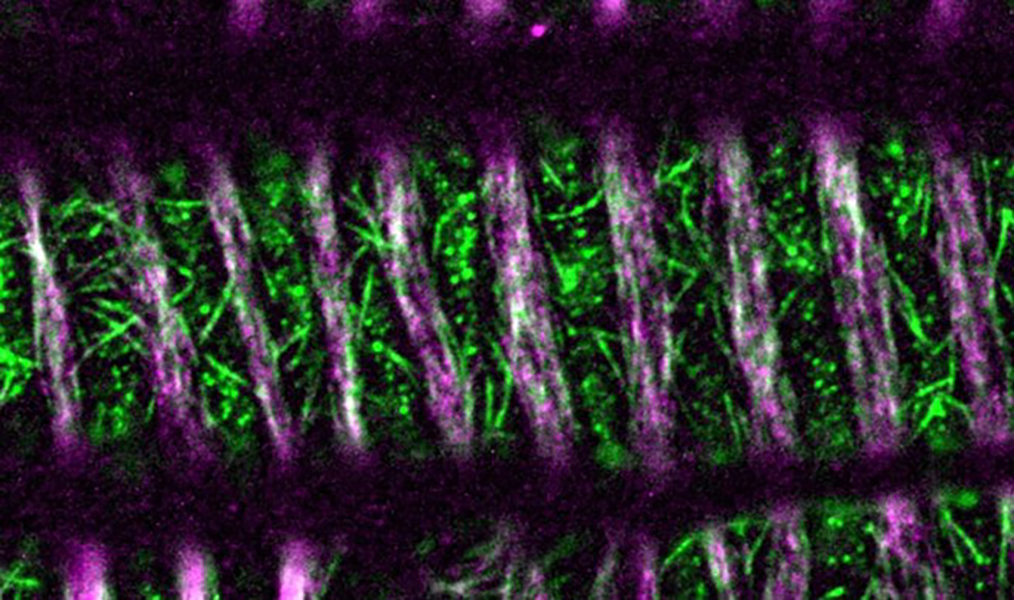
シロイヌナズナの芽生えを透明化処理し、微分干渉顕微鏡を用いて観察した。環状(左)と孔あき状(右)の細胞壁をもった道管が容易に視認される。スケールバーは5マイクロメートルを示す。環状や孔あき状に加え、らせん状や網目状の二次細胞壁をつくる道管もある。