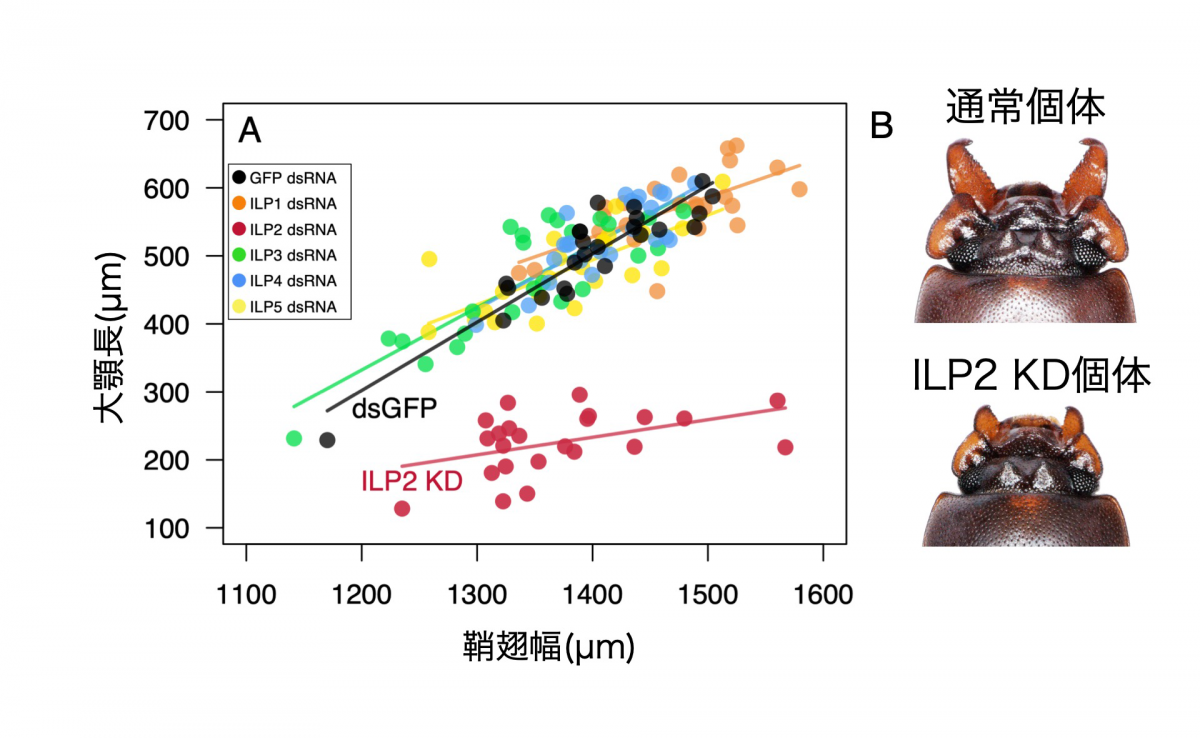
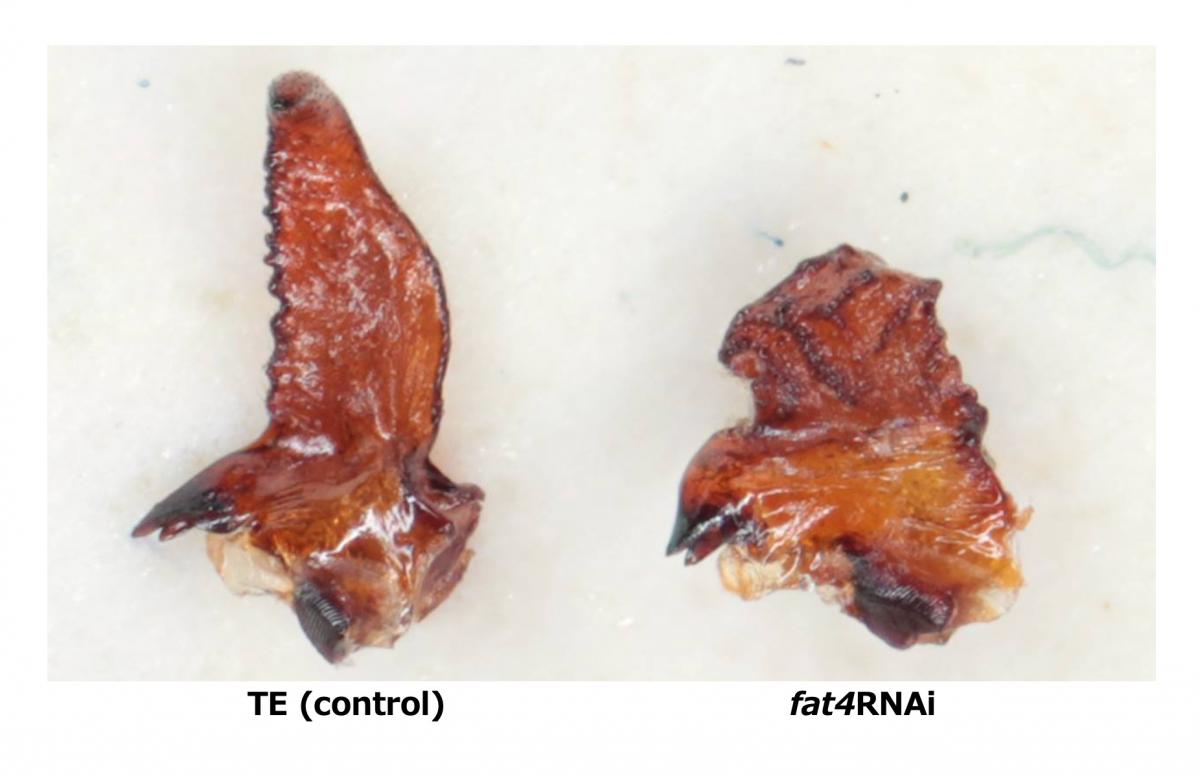
シカやカブトムシに代表されるような動物の武器は、縄張りやメスを巡る闘争で用いられる性選択形質である。繁殖相手の獲得を有利にする形質であるにも関わらず、こうした性的な武器は集団内で大きな変異が存在する(図1)。十分な栄養を得られたオスのみが巨大な武器を発達させる。一方、性的な武器は生存自体に必須ではないため、成長期に栄養が十分得られなかったオスは武器を非常に小さくつくり、分散やスニーキングなど闘争によらない繁殖戦術で子孫を残す。しかしながら、どのような仕組みで武器だけが多様な形態を示すのか、その発生・生理的なメカニズムの多くは未解明である。

図1 オオツノコクヌストモドキの形態変異
左から順に高栄養、中栄養、貧栄養で生育したオス。